作者:Jeffrey P. Tomkins
用分子钟来研究生物进化,这样的的理念对现代生物学已经产生重大影响。所谓分子钟,就是利用不同物种的生物序列(DNA或蛋白质序列)进行比对,以估算进化的速率。
这种估算方法通常采用根据古生物学研究而估算的进化论时间表进行校对(其实这是一种循环论证——译者注)。以估算的时间来校准估算的进化速率,这毫无疑问是一种偏见,除此之外,下面的问题也困扰着该方法的应用。
1. 不同的基因/序列给出了区别极大的进化速率。
2. 在所谓的同源序列上,不同类别的生物展现出不同的进化速率。
3. 即使校对之后,生物类别分化的时间通常与古生物时间表不吻合。
更糟糕的是,由于进化论前提下的分子钟概念,直接与进化论假设所必须的中性模型(neutral model)相关联——然而最近特别是对于全密码子功能(full codon utility)以及全基因组的生化功能的研究,已经彻底否定了该模型的基石。
那么,如果不用这些进化论假设和其漫长时间表去校对分子钟模型,会得出什么结果呢? DNA序列的差异所提供的信息,会不会有助于验证创造论关于起源问题所做出的预测呢?
有意思的是,从非信徒科学家与创造论科学家双方的研究那里,我们已经有了一系列的研究实例。这些研究以经验测量一类生物之内的DNA分子钟(而不是直接根据进化论假设就做出跨生物类别的估算——译者注),并且也不采用进化论漫长时间表做校对。而这些研究所给出的生物存在的历史时间并非数百万年,而是仅在5千到1万年之间。(有趣的是,多个研究表明,当在一个分类单元内进行DNA分子钟研究(而不是直接根据进化论假设就做出跨生物类别的估算——译者注),并且不采用进化论漫长时间表进行校正时,非信徒科学家与创造论科学家都给出的生物存在的历史时间并非数百万年,而是仅在5千到1万年之间。)
这些不同的研究实例在下文会一一给出。但在此之前,让我们先看看与分子钟密切相关的“基因熵”(基因组逐渐衰败——译者注)!
.jpg)
基因熵与基因钟
当突变在减数分裂中发生时,这些突变会被遗传到下一代。以此实际检测一个谱系,就可得出突变的估算速率。事实上科学家已经在多个研究中实际测量了人类基因组中的这个速率,并发现这个速率是每代75到175个 突变。(尾注2-9)
利用这个已知的基因突变速率,多个研究团队已经可以对突变在人类基因组中的随着时间推移的突变的累积做出模型(利用复杂的计算机模拟,并整合了群体遗传学理论中的标准限制因素)(尾注10-16)。他们发现即便处在强大的自然选择之下,依然有高达90%的有害突变无法被剔除。因此,这种突变的累积会到达一个临界点,以至于突变、拷贝错误如此严重而使得人类最终走向灭绝——这个临界点就被称为“误差灾难”(error catastrophe)。(尾注17-18)而生物基因组由于世代的遗传以及因此随时间推移而降解的过程就被称为基因熵。(尾注17-18)
其中引人注意的情况是人类基因缓慢降解的过程,与圣经关于人类寿命递减的过程相互对应——特别是自从大洪水之后的4000-5000年。(尾注15、18-20)
除了大量的模拟研究之外,由智慧设计论的科学家和知名的进化论科学家的研究也显示了人类基因的拷贝错误的累积、或者说突变的积累,无法被自然选择所缓解。(尾注5、21)
这些通过计算机模拟所做出基因熵(基因降解)研究的实验结果发表于多篇研究论文之后(前文涉及的参考文献),其结果以雄辩的方式,更进一步得到另外两项备受瞩目的基因研究的证明——它们基于实际的资料,并得出了根本上相同的结果,甚至与圣经事件的时间框架所吻合。(尾注5、6)
这两项非信仰的研究涉及对人类基因组的蛋白质编码区(外显子)的测序(称为外显子组)(尾注22、23)。它们检测了人类外显子组中出现的占优势的罕见单个核苷酸变异体——一项研究分析了2440个人,另一项研究则检测了6515个人。发现在蛋白质编码的外显子中超过80%的单个核苷酸变异体是有损或有害的(牵涉到遗传疾病)。研究人员将有害突变的不期而至归因于“选择清除的无力”(weak purifying selection)——这实质上便否定了进化论者所声称的自然选择删除有害突变的能力——而这项发现也得到了上述模拟模型的证实。(尾注11、14-16)
对这类型外显子组的罕见变异资料的效益分析是因为蛋白质编码区域比基因组其它部分更不能容忍突变发生。因此这部分就可以给相关研究提供更为可靠的人类种群的基因历史信息。此外,这类型的数据可更方便地被整合到有关已知历史地理信息的人口模型之中。当研究人员着手此类工作后,他们发现世界人口基因的分化是最近才开始爆发的,并主要与“基因熵”过程有关。
其中一份研究报告如此说道:“加速增长的最大可能的时间是5115年之前”(尾注22)。其它研究论文也揭示了相似的时间表——这些就使得人类基因分化的起始时间与创世纪洪水(包括后续的巴别塔人类分散事件)时间极为接近。这里的重点是,很明显,与基因熵进程有关的人类罕见变异的爆发,亦遵从上文所提及的大洪水之后人类寿命急速下跌的模式(参见注释18、20)。
线粒体DNA变异与分子钟
分子钟研究的另外一个重要领域是证实了创造论科学家内森尼尔?金森(Nathaniel Jeanson)关于新近创造的结论。他一直在从事线粒体基因组突变率的研究(尾注24)。我们知道线粒体基因基本上只从母系传承,并且其速率可在家谱中得到准确检测。这就能提供一个谱系钟。这些有关谱系传承的分子钟,如果不经过进化论时间表的假设进行校对,而是利用包括人类在内的某类生物自身的每代时间作为衡量——那么,我们就可以获得更为真实的、没有预设偏见的、针对某类生物的基因历史。通过比较果蝇、蛔虫、水蛭、以及人类等四类生物的实际线粒体分子钟的速率,金森证明了包括人类在内的这些生物被创造的时间点,不会超过1万年!参见图表。
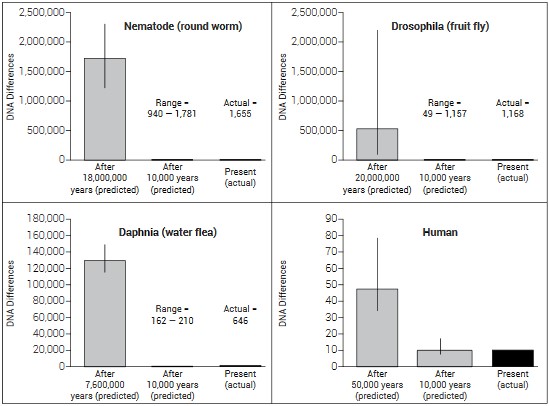 这一组图显示了线粒体DNA多样性的近代起源。每个图的第一、第二、第三个柱形分别代表了进化论假设所做出的多样性的预测(基于其漫长时间表)、1万年时间框架下所做出的多样性预测、以及在这些序列中所实际发现的多样性。黑线代表了95%置信区间。在所有上述四例中,实际数据符合圣经1万年或不到1万年的时间表所给出的预测。数据的计算采用了平均每代时间和实际突变率。上图是经过原作者同意后,根据尾注引文16的资料制作的。 这一组图显示了线粒体DNA多样性的近代起源。每个图的第一、第二、第三个柱形分别代表了进化论假设所做出的多样性的预测(基于其漫长时间表)、1万年时间框架下所做出的多样性预测、以及在这些序列中所实际发现的多样性。黑线代表了95%置信区间。在所有上述四例中,实际数据符合圣经1万年或不到1万年的时间表所给出的预测。数据的计算采用了平均每代时间和实际突变率。上图是经过原作者同意后,根据尾注引文16的资料制作的。 |
另外创造论科学家圣弗德(John Sanford)与卡特(Robert Carter)也研究了人类线粒体DNA的分化——他们统计分析了超过800个不同的序列,并重构了与夏娃原先的线粒体基因组极为接近的序列(尾注18、25)。他们发现人类从夏娃序列中只多出了平均22个突变,尽管有些个体可以多达100个(尾注18)。最近的关于人类线粒体基因的估算是平均每代0.5个(尾注26)。根据这个速率,即便对于突变最多的线粒体序列,圣弗德与卡特亦总结道,“只需要200代(少于6000年)就可以积累到100个变异(尾注18)”。
令人颇感意外是首先提出支持圣经框架时间表的学者正是进化论学家——早在1997年该非信仰研究论文已经发表了,其涉及线粒体突变率的趋势也为新近创造论科学家的研究所观察到。但当时这个论文却在进化论学术领域内几乎没引起关注。(尾注27)这个论文的研究者们如此陈述道:“利用我们实际的速率去校对线粒体分子钟,得到的结论是线粒体DNA MRCA(最近祖先或第一个人类女人)大约在6500年前”。
一年以后,另外一个非信仰科学家就这项研究说道:“进化论者最为关注是更高速率的突变所带来的影响,而不是其产生的原因。例如,研究者已经估算‘线粒体夏娃’(其线粒体基因是所有今天人类的祖先)大约生活于10万到20万年前的非洲。可在新的分子钟的检测下,她只有6000年的历史 。”(尾注28)
该文也注意到,新的关于更高突变速率的发现令线粒体夏娃指向大约6000年前,这项发现甚至还帮助了FBI建立有关法医调查的研究指导原则。现在,利用更多的线粒体基因的数据,金森、卡特和圣弗德毫无争议地证明了之前被进化论学术圈所忽略的研究发现的正确性。
除了线粒体DNA的研究,圣弗德与卡特亦分析了现代人的Y染色体,结果表明现代人与Y染色体亚当的序列相比,平均仅有300个突变(尾注18),因此,他们认为,“即便我们采用通常的Y染色体的突变率(大约每个染色体每代1个突变),获得300个突变也只需300代时间(大约6000年)”。
这与他们之前对于线粒体DNA的研究一样,是对分子钟概念最为直接的运用。其提供的资料完美地印证了圣经记录人类起源的时间框架。
结论
与进化论分子钟的种种错误假设形成鲜明对照(例如进化的确曾经大规模跨类别地发生;使用漫长时间表作为校对;中性模型等等)——在单一分类单元内对生物所做的分子钟研究,表明这些生物基本上存在年限在5000-10000年之间。因此,当我们去掉假想的进化条件,通过实际检测所获得的资料,指向的乃是圣经的时间框架。
【扩展阅读】
参考文献
1. See Tomkins, J.P. and Bergman, J., Evolutionary molecular genetic clocks: a perpetual exercise in futility and failure, J. Creation29(2):26–35, 2015.
2. Nachman, M.W. and Crowell, S.L., Estimate of the mutation rate per nucleotide in humans, Genetics 156:297–304, 2000.
3. Kondrashov, A.S., Direct estimates of human per nucleotide mutation rates at 20 loci causing mendelian diseases, Hum. Mutat.21:12–27, 2003, doi:10.1002/humu.10147.
4. Xue, Y.et al., Human y chromosome base substitution mutation rate measured by direct sequencing in a deep-rooting pedigree, Curr. Biol.19:1453–1457, 2009; doi:10.1016/j.cub.2009.07.032.
5. Lynch, M., Rate, molecular spectrum, and consequences of human mutation, Proc. Natl Acad. Sci. USA 107:961–968, 2010; doi:10.1073/pnas.0912629107.
6. Campbell, C.D. and Eichler, E.E., Properties and rates of germline mutations in humans, Trends Genet.29:575–584, 2013; doi:10.1016/j.tig.2013.04.005.
7. Michaelson, J.J. et al., Whole genome sequencing in autism identifies hot spots for de novo germline mutation, Cell151:1431–1442, 2012; doi:10.1016/j.cell.2012.11.019.
8. Kong, A. et al., Rate of de novo mutations and the importance of father's age to disease risk, Nature 488:471–475, 2012; doi:10.1038/n at u r e11396
9. Roach, J.C. et al., Analysis of genetic inher?itance in a family quartet by whole-genome sequencing, Science 328:636–639, 2010; doi:10.1126/science.1186802.
10. Sanford, J.et al., Mendel’s accountant: A biologically realistic forward-time population genetics program, Scalable Computing 8:147–165, 2007.
11. Sanford , J.et al., Using computer simulation to understand mutation accumulation dynamics and genetic load, Lecture Notes in Computer Science 4488:386–392, 2007.
12. Sanford, J. and Nelson, C., in: Carmen Fuste?( E d.), Studies in Population Genetics, InTech, pp. 117–136, 2012.
13. Brewer, W., Baumgardner, J. and Sanford, J., in: Marks R.J. III (Ed.), Biological Information: New Perspectives, World Scientific Publishing, Hackensack, NJ, pp. 298–311, 2013.
14. Gibson, G. et al., in: Marks R.J. III (Ed.), Biological Information: New Perspectives, World Scientific Publishing, Hackensack, NJ, pp. 232–263, 2013.
15. Nelson, C. and Sanford, J., in: Biological Information: New Perspectives, Marks R.J. III (Ed.), World Scientific Publishing, Hackensack, NJ, pp. 338–368, 2013.
16. Sanford, J., Baumgardner, J. and Brewer, W., Marks R.J. III (Ed.), Biological Information: New Perspectives, World Scientific Publishing, Hackensack, NJ, pp. 264–297, 2013.
17. Sanford, J., Genetic Entropy and the Mystery of the Genome, 3rd edn, FMS Publications, 2010.
18. Sanford, J. and Carter, R., In light of genetics... Adam, Eve, and the Creation/Fall, Christian. Apologetics. J. 12:51–98, 2014.
19. Osgood, J., The date of Noah’s Flood, Creation 4:10–13, 1981.
20. Sanford, J., Pamplin, J. and Rupe, C., Genetic entropy recorded in the Bible?, www.kolbecenter.org./wp-content/uploads/2014/07/Genetic-Entropy-Recorded-in-the-Bible.pdf%3E, 2014.
21. Crow, J.F., The high spontaneous mutation rate: Is it a health risk? Proc. Natl Acad. Sci. USA 94: 8380–8386, 1997.
22. Tennessen, J.A.et al., Evolution and functional impact of rare coding variation from deep sequencing of human exomes, Science 337:64–69, 2012; doi:10.1126/science.1219240.
23. Fu, W.et al., Analysis of 6,515 exomes reveals the recent origin of most human protein-coding variants, Nature 493:216–220, 2013;doi:10.1038/n at u r e1169 0.
24. Jeanson, N., Recent, functionally diverse origin for mitochondrial genes from ~2700 metazoan species, Answers Research J.6:467–501, 2013.
25. Carter, R.W., Mitochondrial diversity within modern human populations, Nucleic Acids Res. 35:3039–3045, 2007; doi:10.1093/nar/gkm207.
26. Madrigal, L.et al., High mitochondrial mutation rates estimated from deep-rooting Costa Rican pedigrees, Am. J. Phys. Anthropol.148:327–333, 2012; doi:10.1002/ajpa.22052.
27. Parsons, T.J.et al., A high observed substitution rate in the human mitochondrial DNA control region, Nat. Genet.15:363–368, 1997; doi:10.1038/ng0497-363.
28. Gibbons, A., Calibrating the mitochondrial clock, Science 279:28–29, 1998.
原文见:国际创造论事工 www.creation.com
